A imunidade adquirida pela infecção natural de um vírus é melhor que a imunidade adquirida por uma vacina?
Aprenda tudo que você precisa saber sobre imunologia e descubra porque tomamos várias doses da vacina contra a Covid-19.

Direto ao ponto: se a variante do vírus não tiver adquirido muitas mutações após o desenvolvimento da vacina que está sendo aplicada, então ambas as imunidades induzidas pelas vacina e infecção natural são equivalentes. No entanto, vale ressaltar que precisamos primeiro sobreviver uma infecção natural para poder adquirir imunidade ao vírus; ou seja, a infecção natural sempre te coloca em risco (além de poder deixar sequelas graves). Já as vacinas aprovadas pelas agências reguladoras sempre te proporcionam algum nível de imunidade sem colocar sua vida em risco. O problema surge quando uma vacina fica defasada em relação à variante do vírus que está em circulação, pois nesta ocasião ela deixa de proporcionar uma imunidade adequada. Para entender os diferentes níveis de imunidade é necessário primeiramente entender como os anticorpos e o sistema imune adaptativo atuam para nos proteger de uma reinfecção pelo vírus.
Se você não tem curiosidade em entender os pormenores do nosso sistema imunológico, leia apenas o primeiro tópico, que aborda a imunidade humoral, e depois vá direto para o penúltimo tópico, que aborda as vacinas.
Tabela de conteúdo:
➡️Como os anticorpos nos protegem?
➡️Quais os diferentes tipos de anticorpos e por que são importantes?
➡️Como o sistema imune adaptativo consegue se adaptar a qualquer antígeno tão eficientemente?
➡️Como os diferentes tipos de linfócitos B virgens atuam na resposta imune adquirida?
➡️Como que o sistema imune adaptativo diferencia aquilo que é nosso (autoantígeno) daquilo que é invasor (antígeno)?
➡️Por que precisamos de células dendríticas, linfócitos T e linfócitos B para desencadear a resposta humoral?
➡️O que acontece quando somos vacinados ou infectados por um vírus?
➡️Dados recentes sobre a segurança das vacinas
Como os anticorpos nos protegem?
A imunidade mediada por anticorpos é conhecida como imunidade humoral e ela nos protege por meio de três processos: (1) neutralização; (2) opsonização; e (3) ativação do complemento.
Destes, a neutralização é a mais relevante para a proteção contra vírus, de maneira que focarei apenas nela. Para penetrar na célula, o vírus precisa se ligar a moléculas específicas (receptores) na superfície da célula-alvo (Figura 1A).
Já os anticorpos ligam-se especificamente ao vírus e impedem que este se ligue às células-alvo (Figura1D); diz-se que os anticorpos neutralizam o vírus. Sem conseguir infectar as células-alvo para se multiplicar, o vírus é facilmente eliminado por células de defesa.

As células responsáveis pela produção de anticorpos são os linfócitos B (ou células B), que no estado inativado (células B virgens) produzem anticorpos inespecíficos na superfície de sua membrana plasmática.
Estes anticorpos, chamados de imunoglobulinas de superfície, podem se ligar a uma variedade de estruturas químicas, incluindo partículas virais inteiras (Figura 2A).
Quando isso acontece, a partícula viral é internalizada no linfócito B e degradada (Figura 2B).
Os fragmentos de proteínas do vírus, chamados antígenos, são então encaminhados para a superfície do linfócito B virgem para que sejam apresentados aos linfócitos T auxiliares, também chamados de células T auxiliares (Figura 2C).
Se os linfócitos T auxiliares reconhecerem o antígeno apresentado pelo linfócito B virgem, haverá a liberação de citocinas (moléculas estimuladoras) que induzirão o linfócito B a se transformar em uma célula B ativada, chamada célula plasmática.
A célula plasmática é capaz de produzir e secretar anticorpos específicos contra a mesma proteína do vírus que foi reconhecida pelo linfócito T auxiliar (Figura 2D); por fim, os anticorpos específicos atuarão na neutralização do vírus.
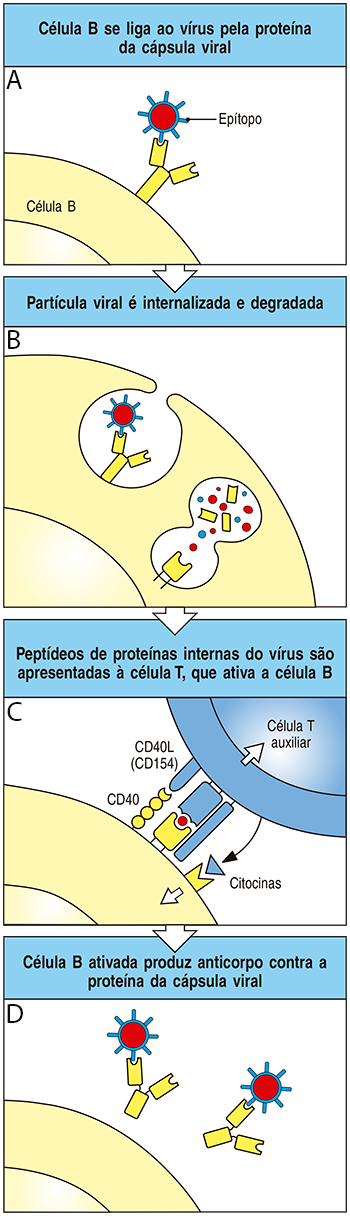
Você pode estar se perguntando: por que é necessário que a produção de anticorpos específicos ocorra somente após a apresentação do antígeno viral aos linfócitos T auxiliares? Isso ocorre porque a imunoglobulina de superfície dos linfócitos B virgens é um anticorpo inespecífico que possui baixa afinidade a antígenos estruturalmente similares, podendo inclusive se ligar acidentalmente a moléculas nativas, ou autoantígenos.
Dessa forma, podemos imaginar o linfócito B virgem como um ‘papiloscopista’ que coleta ‘impressões digitais’ pelo nosso corpo por meio de amostragem. Nessa analogia, os linfócitos T auxiliares atuariam como ‘agentes da polícia’, que analisam os dados coletados para averiguar se pertencem a algum suspeito envolvido em crimes; neste caso, algum vírus.
Caso a impressão digital pertença a um cidadão de bem (autoantígeno), o policial (linfócitos T auxiliar) ignora o papiloscopista (linfócito B virgem). Caso pertença, o policial emite um ‘mandado de prisão’ para capturar o suspeito, que no caso dos linfócitos T auxiliares seria equivalente ao papel desempenhado pelas citocinas (Figura 2C).
Uma vez informado do mandado de prisão (citocinas), o papiloscopista (linfócito B virgem) é ativado para neutralizar o suspeito usando ‘algemas’ (sim, papiloscopistas podem prender suspeitos); seguindo essa lógica, as algemas, acrescidas do mandado de prisão, representariam os anticorpos específicos produzidos pela célula plasmática (o papiloscopista encarregado de prender o suspeito).
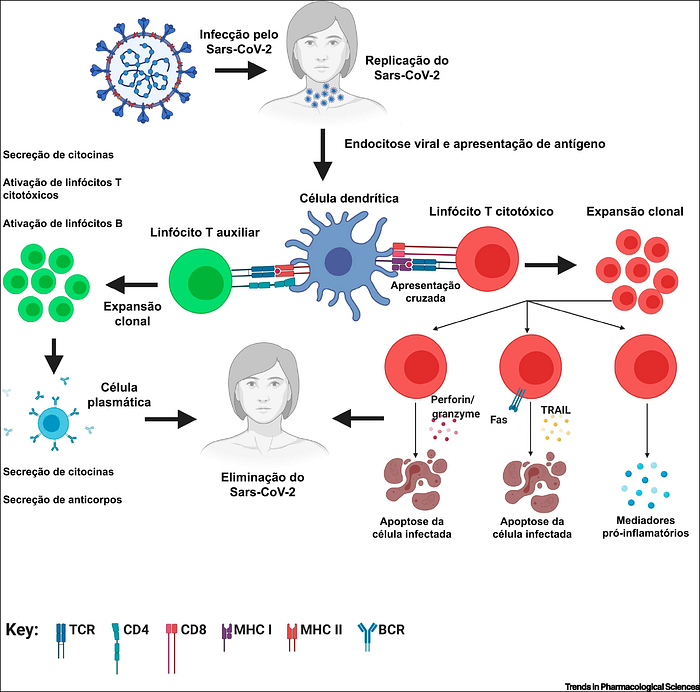
Para finalizar nossa analogia, é preciso entender como os linfócitos T auxiliares (agentes da polícia) conseguem diferenciar se um antígeno (impressão digital) pertence, ou não, a algum vírus (alguém suspeito de cometer crime). Aí entra o trabalho das células dendríticas (Figura 3).
Essas células atuam como verdadeiros “detetives”: elas patrulham todo o nosso corpo em busca de pistas e sempre que encontram alguma impressão digital suspeita (antígeno), a apresentam para os linfócitos T virgens (policiais em formação).
Ao serem “informados”, os linfócitos T virgens se diferenciam em linfócitos T auxiliares (agentes da polícia) ou linfócitos T citotóxicos, ambos capazes de reconhecer especificamente o antígeno que lhes foi apresentado pela célula dendrítica; a definição entre se diferenciar em auxiliar ou citotóxico depende do tipo de correceptor que eles carregam (CD4 ou CD8, respectivamente).
Uma vez diferenciados, os linfócitos T auxiliares serão capazes de ativar os linfócitos B virgens que apresentarem o mesmo antígeno o qual foram ativados para reconhecer. Já os linfócitos T citotóxico serão capazes de destruir as células do nosso corpo que foram infectadas pelo vírus detentor do antígeno, evitando assim que este se multiplique e infecte ainda mais células (Figura 3).
Isso acontece porque todas as células do nosso corpo estão equipadas com moléculas do Complexo Principal de Histocompatibilidade (MHC) de classe I, que as permitem apresentar aos linfócitos T citotóxicos os antígenos de um vírus que as tenha infectado.
Portanto, enquanto as células plasmáticas são responsáveis por secretar anticorpos pra evitar que o vírus penetre nas células-alvo, os linfócitos T citotóxicos são responsáveis por eliminar as células que já foram infectadas pelo vírus.
É importante ressaltar que a imunidade adaptativa, composta pela imunidade humoral e imunidade mediada por células, depende da participação de todos os personagens mencionados acima, começando pelas células dendríticas (veja o vídeo acima).
Os linfócitos B só produzem anticorpos específicos para um antígeno após receberem um sinal positivo dos linfócitos T auxiliares, e estes, por sua vez, só são capazes de ativar os linfócitos B se tiverem sido previamente ativados por células dendríticas.
Portanto, as células dendríticas formam o elo entre a imunidade inata (aquela que nasce conosco) e imunidade adaptativa (aquela que adquirimos após o contato inicial com um vírus).
O motivo disso é que essas células acumulam uma coleção de receptores especializados em reconhecer todo tipo de moléculas invasoras. Essa coleção de receptores vem sendo constantemente aperfeiçoada durante a evolução do sistema imune inato, de forma a permanecer sempre atualizada.
No entanto, a origem dessa coleção tão abrangente e eficiente de receptores ainda é incerta, não sabemos se a adquirimos através da evolução dos nossos próprios genes ou se seria uma herança inusitada de infecções ancestrais. A resposta mais provável é que ambas as fontes contribuem para essa robustez, mas com pesos diferentes.
Você pode estar se perguntando “mas se as células dendríticas já possuem todos esses receptores para reconhecer moléculas invasoras, então por que elas mesmas não produzem os anticorpos? Por que é preciso ativar primeiro linfócitos T para que depois estes ativem os linfócitos B? Não seria mais inteligente (e eficiente) fazer tudo de uma vez?”.
Nos tópicos seguintes veremos o que são anticorpos, como são refinados, e qual a sua relação com as doenças autoimunes. Esses conceitos serão importantes para entender por que o sistema imune criou um grupo de células especializadas em produzir os anticorpos, ao invés de delegar essa função para as células dendríticas.
No entanto, pode-se pular essas informações e ir direto ao tópico relativo às vacinas sem grandes prejuízos para o entendimento.
Quais os diferentes tipos de anticorpos e por que são importantes?
O processo de transformação dos linfócitos B virgens em células plasmáticas (secretoras de anticorpos) passa por duas etapas: a maturação de afinidade e a troca de classe dos anticorpos.
De maneira simplificada, a maturação de afinidade é um processo de mutação por meio do qual um anticorpo inespecífico do linfócito B virgem é repetidamente refinado de maneira a aumentar sua afinidade e especificidade ao antígeno viral.
Lembrando que afinidade diz respeito ao quão forte o anticorpo se ligará ao antígeno; uma ligação de alta afinidade indica que anticorpo permanecerá ligado ao antígeno por mais tempo. Já a especificidade diz respeito a capacidade do anticorpo de se ligar apenas àquele antígeno; uma ligação com alta especificidade implica dizer que o anticorpo não irá reconhecer outros tipos moléculas.
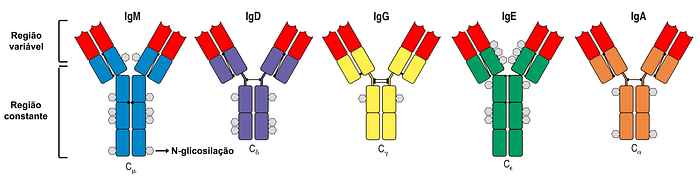
Já a troca de classe é o processo pelo qual um tipo de anticorpo, ou imunoglobulina (Ig), é convertido em outro tipo. Aos diferentes tipos de imunoglobulinas damos o nome de isotipos.
Existem cinco principais isotipos de imunoglobulinas, denominados IgM, IgD, IgG, IgE e IgA; todos eles podem atuar tanto como receptores de antígenos ancorados à membrana de linfócitos B (como a imunoglobulina de superfície; Figura 2A) quanto como anticorpos secretados (solúveis) na corrente sanguínea (Figura 2D).
Cada isotipo possui duas regiões: a região variável (V), a qual é responsável por se ligar aos antígenos (Figura 4, em vermelho); e a região constante (C), ou invariável, a qual é responsável pelas diferentes formas como os anticorpos podem atuar na sinalização e remoção de antígenos (Figura 4, em diferentes cores).
Enquanto a região V é suficiente para neutralizar um vírus, isto é, ligar-se fortemente a ele para impedir sua infecção na célula-alvo (Figura 1D), a região C é essencial para recrutar a ajuda de outras células e moléculas para destruir e remover o vírus ao qual o anticorpo se ligou.
Por exemplo, a região C de IgG recruta macrófagos e neutrófilos, enquanto a região C de IgE recruta mastócitos, basófilos e eosinófilos.
Já a região C de IgA permite que esse anticorpo forme dímeros (grupos de duas moléculas) e seja secretado nas mucosas que revestem o trato respiratório e digestivo, bem como nas lágrimas e no leite, enquanto a região C de IgM permite a formação de pentâmeros e a rápida ativação do complemento (uma outra forma pela qual os anticorpos nos protegem, diferente da neutralização).

Por fim, IgD possui uma região C flexível que desfavorece ligação a antígenos monovalentes (que possuem apenas uma região para anticorpos se ligarem), favorecendo assim a detecção de polímeros e complexos multivalentes.
Portanto, a troca de classe permite que a região V de uma imunoglobulina possa se associar a qualquer região C dos isotipos de imunoglobulina.
Isso permite que um linfócito B produza uma variedade de anticorpos que compartilham a mesma especificidade e afinidade pelo antígeno, porém estejam melhor adaptados para atuar em diferentes funções e tecidos do nosso corpo.
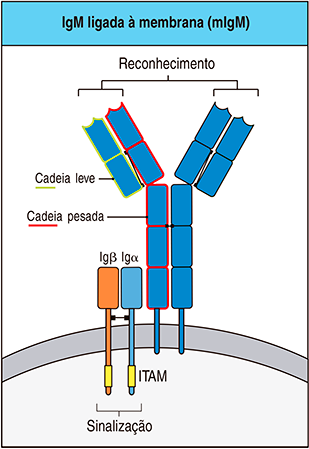
Antes de serem ativados pelos linfócitos T auxiliares, os linfócitos B virgens possuem apenas imunoglobulinas de superfície (sIg) ancoradas às suas membranas plasmáticas, que atuam como receptores inespecíficos de antígenos.
Por este motivo, as imunoglobulinas de superfície dos linfócitos B virgens fazem parte de um grupo maior de proteínas chamado Receptores do Linfócito B (cuja sigla em inglês é BCR, em referência a “B Cell Receptors”); este grupo maior inclui também as imunoglobulinas de membrana (mIg), que atuam como receptores específicos de antígenos (Figura 5).
Enquanto as mIgs estão presentes em linfócitos B ativados e podem pertencer a qualquer classe de isotipo, as sIgs, por sua vez, pertencem unicamente a duas classes de isotipos: IgM e IgD; por motivo de clareza, chamamos os dois isotipos das sIgs de IgM de superfície e IgD de superfície.
Os linfócitos B virgens apresentam tanto IgM quanto IgD de superfície ancorados em suas membranas, mas podemos dividí-los em dois grupos: linfócitos ricos em IgM e linfócitos ricos em IgD. Enquanto o primeiro grupo apresenta muito mais IgM do que IgD de superfície (são ditos IgM+/IgD), no segundo grupo essa relação é invertida (são ditos IgD+/IgM).
Os linfócitos B virgens que mencionei anteriormente, os quais são ativados por linfócitos T auxiliares para secretar anticorpos de maior afinidade e especificidade ao antígeno, são chamados linfócitos B foliculares e pertencem ao grupo de linfócitos IgD+/IgM (ricos em IgD); ou seja, os anticorpos produzidos pela ativação de linfócitos IgD+/IgM sofrem maturação de afinidade e troca de classe.
Já os linfócitos IgM+/IgD (ricos em IgM) não precisam ser ativados por linfócitos T auxiliares para secretar anticorpos solúveis, basta que eles se liguem ao antígeno; exemplos deste grupo são os linfócitos B1 e linfócitos da zona marginal (MZ).
No entanto, os anticorpos secretados por esses linfócitos se restringem principalmente ao isotipo IgM e apresentam baixa afinidade e especificidade ao antígeno.
Embora esses anticorpos possam ser menos eficientes e até mesmo prejudicais aos nossos tecidos, eles se fazem necessários por serem produzidos rapidamente, podendo assim atuar de forma imediata na neutralização do vírus.
Como veremos mais adiante, essa diferença entre linfócitos IgM+/IgD e IgD+/IgM é extremamente relevante para entender como o sistema imune adaptativo diferencia aquilo que é nosso (autoantígeno) daquilo que é invasor (antígeno).
Além disso, a prevalência do isotipo IgM ou IgD na superfície celular determina a velocidade e o tipo de resposta desencadeada pelos linfócitos B virgens, o que tem importantes implicações para a imunidade induzida por vacinas.
Como o sistema imune adaptativo consegue se adaptar a qualquer antígeno tão eficientemente?
É importante salientar que embora os linfócitos B virgens possuam mais de um isotipo de imunoglobulina, cada linfócito produz individualmente apenas uma região V para essas imunoglobulinas.
Ou seja, a mesma região V estará presente nos diferentes isotipos de imunoglobulinas de um mesmo linfócito B, mas cada linfócito produzirá uma região V diferente em relação ao restante dos linfócitos.
O mesmo vale para os tipos celulares que se diferenciam a partir dos linfócitos B virgens, como as células plasmáticas.
Esse fenômeno é a base para a Teoria da Seleção Clonal (agraciada com o Prêmio Nobel), que afirma que, para um mesmo linfócito, haverá apenas a produção de anticorpos com a mesma região V.
Essa teoria explica como o sistema imune adaptativo consegue responder a diferentes tipos de infecção tão eficientemente, mesmo que partindo de um único linfócito produzindo anticorpos com baixa afinidade e especificidade ao antígeno (também chamado de polireatividade e poliespecificidade, respectivamente).
Um anticorpo de baixa afinidade é chamado de polireativo porque diferentes antígenos com formatos parecidos podem se ligar ao mesmo sítio de ligação desse anticorpo.
Já um anticorpo poliespecífico possui uma região V que acomoda mais de um sítio de ligação, podendo se ligar a antígenos com formatos diferentes.
Perceba que cada sítio de ligação diferente presente em uma região V de um anticorpo poliespecífico pode, portanto, também apresentar polireatividade.
Dessa forma, ao produzir inicialmente anticorpos polireativos e poliespecíficos, nosso sistema imune tenta primeiro preencher toda a gama de possíveis formatos que um antígeno pode adotar.
Uma vez selecionado o linfócito produzindo um anticorpo capaz de reconhecer o antígeno invasor, o sistema imune prioriza a geração de vários clones desse linfócito e inicia um esforço para refinar a afinidade e especificidade do anticorpo por ele produzido (veja o vídeo acima).
Mas afinal de contas, de quantos clones precisamos para conseguir detectar todos os potenciais formatos de antígenos, assumindo que os anticorpos sejam polireativos e poliespecíficos?
Menos do que se imagina — dados teóricos e experimentais parecem convergir para algo entre 10 e 20 milhões de clones [74–76]. Se não fosse pela polireatividade e poliespecíficidade, esse valor seria pelo menos cinco ordens de grandeza maior, entre 10¹²–10¹⁵ clones [77].
Portanto, existe uma grande vantagem evolutiva para os linfócitos B reconhecerem inicialmente o formato de antígenos de forma polireativa e poliespecífica, para só depois se especializarem na detecção do antígeno.
No entanto, essa flexibilidade também abre brecha para que os linfócitos B sejam reativos a moléculas do próprio hospedeiro, de forma que a natureza precisou evoluir outros mecanismos para controlar o reconhecimento dos autoantígenos.
Como os diferentes tipos de linfócitos B virgens atuam na resposta imune adquirida?
Para que linfócitos ricos em IgD (IgD+/IgM) sejam ativados por linfócitos T auxiliares e refinem seus anticorpos após interagir com um antígeno, é necessário que eles primeiro migrem para locais específicos do nosso corpo, os folículos linfoides [2].
Uma vez ativados pelos linfócitos T, formam centros de germinação (CG) onde ocorrem a maturação de afinidade e troca de classe de seus anticorpos. Não é surpresa que a conjunção dessas etapas requera um certo tempo para ser concluída.
No entanto, tempo é algo precioso quando estamos sendo atacados por um vírus; dessa forma, nosso corpo também desenvolveu um mecanismo para produzir anticorpos a curto prazo (a natureza é sábia)!
Enquanto os linfócitos B foliculares (IgD+/IgM) estão a se diferenciar em células plasmáticas nos CGs, os linfócitos B1 e linfócitos da zona marginal (ambos do tipo IgM+/IgD) se diferenciam rapidamente em plasmoblastos sem precisar da ajuda dos linfócitos T [58].
O plasmoblasto é um estágio anterior ao da célula plasmática, mas que já é capaz de secretar grandes quantidade de IgM de baixa afinidade ao antígeno [63].
A escolha em secretar o isotipo IgM não é por acaso. Estes são sempre os primeiros anticorpos a serem secretados durante uma resposta imune humoral pois são produzidos sem precisar passar pelo processo de troca de classe [53].
Secretar IgM primeiro também é vantajoso por causa das características de sua região C: embora as moléculas de IgM secretadas por plasmoblastos sejam anticorpos de menor afinidade, a região C deste isotipo permite com que ele forme pentâmeros (agrupamentos de cinco moléculas).
Um pentâmero de IgM possui mais sítios de ligação (dez) que um monômero (dois), de maneira que isso compensa a afinidade relativamente baixa ao antígeno ao permitir a ligação a múltiplos pontos (quanto maior o número de interações entre o anticorpo e o antígeno, maior é a avidez do anticorpo).
Por exemplo, um pentâmero pode se ligar a dez proteínas de spike do coronavírus, aumentando em cinco vezes a força de ligação ao vírus em comparação a um monômero (duas proteínas de spike).
Além de IgM, os plasmoblastos podem vir a secretar IgG por meio da troca de classe, mas somente se forem estimulados, através de citocinas, por células dendríticas que estiveram nas proximidades.
Também podem sofrer algumas poucas mutações na região V de seus anticorpos em um processo similar à maturação de afinidade, mas intermediado por receptores do tipo Toll presentes em suas membranas e sem a necessidade de formar centros germinativos.
Os receptores do tipo Toll fazem parte do nosso sistema imune inato e são apenas um dos muitos receptores de moléculas invasoras que também estão presentes em células dendrítica.
Quando o processo de mutação é intermediado por receptores do tipo Toll, em contraposição à maturação de afinidade desencadeada pelos linfócfitos T auxiliares, o linfócito IgM+/IgD equipara as quantidades de IgM e IgD de membrana que ele produz e temos a formação de células B de memória do tipo IgM (embora elas sejam, na prática, IgM/IgD), também conhecidas como células B de memória naturais.
Na prática, essas células atuam da mesma forma que os linfócitos B1 e linfócitos da zona marginal, com a diferença que são mais eficientes e também circulam por regiões onde os outros dois estão menos presentes.
Como que o sistema imune adaptativo diferencia aquilo que é nosso (autoantígeno) daquilo que é invasor (antígeno)?
Já vimos que células de defesa do sistema imune inato (aquele que nasce conosco), como as células dendríticas, possuem uma coleção de receptores que lhes permitem identificar moléculas invasoras em meio a todas as outras.
Mas como se dá esse processo nas células do sistema imune adaptativo?
Uma vez que o contato inicial do linfócito B com moléculas invasoras é intermediado pelas imunoglobulinas de superfície em sua membrana, este deve primeiro evitar produzir imunoglobulinas que possuam alta afinidade aos autoantígenos, caso contrário desenvolver-se-ia uma reação infamatória contra o próprio corpo.
Já em um segundo momento, ao se adaptar a um novo invasor, o anticorpo produzido pelo linfócito B sofre diversas mutações randômicas, de maneira que este também deve evitar que essas mutações incorram em uma “adaptação” aos autoantígenos.
Portanto, em ambas essas situações deve haver um mecanismo de controle que impeça o surgimento de células autoreativas. De fato, hoje sabemos que os linfócitos B passam por uma espécie de treinamento para evitar reconhecer moléculas do próprio hospedeiro.
Esse treinamento acontece na medula óssea e no centro dos folículos linfoides e utiliza uma lógica bastante simples: aquilo que é nosso estará presente em maior quantidade do que aquilo que é invasor, de maneira que células superestimuladas durante o treinamento são consideradas autoreativas e, portanto, “reprovam” [3].
Embora a lógica seja simples, o ato de treinar nosso sistema imune para quele saiba diferenciar entre antígenos e autoantígenos é um dos processos mais complexos e bem regulados do nosso corpo, possuindo várias etapas de verificação.
Ainda assim, está suscetível a erros, cuja consequência mais óbvia é o desenvolvimento de doenças autoimunes como o Lúpus Eritomatoso Sistêmico.
Da mesma forma, é comum o relato de pessoas que desenvolveram uma doença autoimune depois de alguma infecção por agente patogênico; isso acontece quando os mecanismos de controle não conseguem evitar que alguns linfócitos se tornem autoreativos após a maturação de afinidade de seus anticorpos.
O desenvolvimento dos linfócitos B inicia-se na medula óssea a partir de células-tronco, mas o treinamento mencionado acima inicia-se somente a partir da última etapa antes de se transformarem em células B maduras (ou, como conhecemos, linfócitos B virgens).
Durante essa etapa de treinamento, os linfócitos são chamados de células B imaturas e possuem apenas IgM de superfície em suas membranas. Como já fora mencionado, as imunoglobulinas de superfície são inespecíficas e possuem baixa afinidade a antígenos; são, portanto, mais reativas.
Para garantir que as células polireativas não sejam ativadas a todo momento, mas somente quando encontram grandes quantidades de antígeno, a resposta celular induzida pelas imunoglobulinas de superfície só é efetiva quando estas se somam.
Alternativamente, menos imunoglobulinas precisam estar engajadas na ligação ao antígeno quando estas participam em ligações cruzadas.
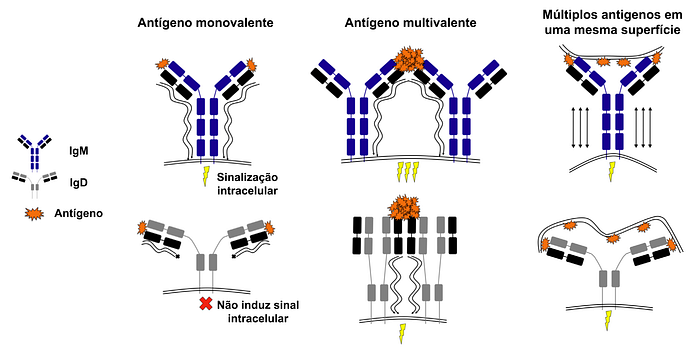
A ligação cruzada ocorre quando um antígeno se liga a duas ou mais imunoglobulinas de superfície na membrana do linfócito (ver na Figura 6); em geral, a ligação cruzada é desencadeada por um antígeno multivalente e causa um acúmulo das imunoglobulinas de superfície em um mesmo local da membrana.
Esse acúmulo proporciona sinergia entre os receptores e amplifica os sinais que eles emitem para o interior da célula, diminuindo a necessidade de grandes quantidades de antígeno para desencadear uma resposta celular efetiva.

No entanto, o isotipos IgM e IgD possuem diferentes sensibilidades às ligações cruzadas e isso está intimamente relacionado à capacidade dos linfócitos em detectar autoantígenos.
A ligação de um antígeno monovalente à IgM de superfície gera sinais normais, enquanto antígenos multivalentes geram um sinal exacerbado que induz a célula na entrar em apoptose (morte celular programada).
Em contrapartida, o isotipo IgD possui uma região flexível que aumenta a entropia das regiões variáveis desta imunoglobulina (Figura 7), fazendo com que seja necessária sua ligação a antígenos multivalentes para conseguir estabilizar a interação entre antígeno e anticorpo (o que gera um aumento significativo da entalpia); em paralelo, a entropia intrínseca do isotipo IgD desfavorece sua ligação a antígenos monovalentes, que são incapazes de gerar um aumento significativo da entalpia (Figura 7).
Dessa forma, toda vez os linfócitos precisam ser mais sensíveis a presença de um antígeno ou autoantígeno, estes produzem quase que exclusivamente IgM de superfície; já quando precisam ser mais toleráveis às concentrações do antígeno ou autoantígeno, aumentam a produção de IgD de superfície.
Não por acaso, os dois momentos onde os linfócitos B produzem exclusivamente IgM de superfície acontecem quando estes precisam eliminar células autoreativas: antes de se diferenciarem em linfócitos B virgens e durante a maturação de afinidade dos anticorpos.
Como o treinamento dos linfócitos B acontece em locais onde antígenos tem seu acesso controlado por macrófagos e células dendríticas, há sempre uma presença muito maior de autoantígenos nesses locais.
Logo, clones cujas imunoglobulinas IgM induzam uma resposta celular efetiva durante o treinamento, seja por ligação cruzada ou pela ativação de muitos receptores, deverão primeiro modificar a região V de seus anticorpos e, se ainda assim continuarem induzindo uma reposta efetiva, deverão entrar em apoptose.
Portanto, linfócitos que produzem IgM altamente autoreativa são eliminados durante o treinamento, ficando apenas aqueles que tem menor chance de causar uma reação autoimune.
Infelizmente essa eliminação de células autoreativas também cria brechas para que agentes patogênicos produzam moléculas invasoras “camufladas”; ou seja, que se assemelham as nossas próprias moléculas e, portanto, são capazes de escapar o sistema imune.
Como isso é claramente perigoso para nós, o sistema imune precisou desenvolver um mecanismo de controle adicional.
Embora linfócitos B altamente autoreativos sejam eliminados durante o treinamento, é permitido que células B fracamente autoreativas se desenvolvam em linfócitos B virgens.
A lógica por trás disso é garantir que os linfócitos B produzam anticorpos que detectem todos os possíveis formatos de antígenos, mesmo que isso implique em manter linfócitos autoreativos potencialmente perigosos para nós mesmos.
No entanto, como forma de diminuir o risco potencial, esses linfócitos B não são inteiramente funcionais e possuem diversas de suas capacidades inativadas, incluindo a capacidade de internalizar e apresentar antígenos (Figura 2B e 2C).
Dessa forma, não podem ser ativados pelos linfócitos T auxiliares e não são capazes de secretar anticorpos. Dizemos que esses linfócitos estão em um estado de anergia.
Os sinais celulares que fazem com que um linfócito B entre e permaneça em estado de anergia são desencadeados principalmente pela IgM de superfície durante as fases de treinamento mencionadas anteriormente.
Para amadurecem em linfócitos B virgens (anérgicos ou não), as células B imaturas precisam produzir IgD de superfície. No entanto, a quantidade de IgD que será produzida depende se a célula é anérgica ou funcional.
Células B anérgicas produzem a maior quantidade de IgD e se diferenciam em linfócitos B foliculares anérgicos (do tipo IgD+/IgM); enquanto células B funcionais podem produzir pouco ou mais IgD, se diferenciando em linfócitos da zona marginal (do tipo IgM+/IgD) e linfócitos B foliculares (do tipo IgD+/IgM), respectivamente. Cerca de 30–50% dos nossos linfócitos B foliculares são anérgicos.
Duas perguntas surgem: (1) por que mantemos linfócitos anérgicos (potencialmente autoreativos) em nosso corpo se estes não são capazes de secretar anticorpos; e (2) por que linfócitos anérgicos precisam produzir mais IgD?
No caso da primeira pergunta, o motivo é que esses linfócitos podem se tornar úteis caso sejamos infectados por um agente patogênico que utilize moléculas similares às nossas próprias. Quando isso acontece, nosso sistema imune falha em conter a infecção e há um aumento significativo no número de moléculas invasoras “camufladas”.
Embora essas moléculas camufladas possam ser detectadas em baixa quantidade pelos anticorpos dos linfócitos B anérgicos, estas precisam estar em grandes quantidades para serem concomitantemente detectadas pelos receptores do tipo Toll que também estão presentes nestes linfócitos.
Como já fora mencionado, os receptores do tipo Toll fazem parte do nosso sistema imune inato e são apenas um dos muitos receptores que também estão presentes em células dendrítica.
A dupla detecção de antígenos pelas imunoglobulinas de superfície e receptores do tipo Toll envia um sinal intracelular que permite aos linfócitos B anérgicos se tornarem funcionais novamente; chamamos esse processo de redenção clonal dos linfócitos anérgicos.
Durante a redenção clonal, os anticorpos passam pelo processo de maturação de afinidade e são selecionados de maneira a aumentar a afinidade pelas moléculas invasoras camufladas e, ao mesmo tempo, diminuir a afinidade ao(s) autoantígeno(s), o que acontece com ajuda de linfócitos T reguladores.
Linfócitos T reguladores são linfócitos T auxiliares autoreativos que se tornaram anérgicos e adquiriram a capacidade de inibir a proliferação de linfócitos B autoreativos nos centros germinativos.
É importante perceber que o mecanismo de redenção clonal é acionado apenas em casos extremos: nosso sistema imune mantém os linfócitos B potencialmente autoreativos em um estado não funcional (anergia), mas permite a ativação destes quando está perdendo a batalha para um agente patogênico.
Ou seja, há situações onde vale a pena o risco de causar dano a si mesmo, desde que para eliminar um invasor sabidamente mais perigoso (já falei que a natureza é sábia?).
No caso da segunda pergunta, o motivo é que IgD é menos reativo que IgM (desfavorece ligação a autoantígenos monovalentes) e produz, uma vez que é efetivamente estimulado, sinais menos anérgicos que IgM.
Ou seja, IgD diminui a ligação de linfócito anérgicos aos autoantígenos monovalentes e facilita a ativação da redenção clonal, caso esta venha a ser necessária.
Em resumo, as células B imaturas altamente autoreativas são eliminadas durante as fases de treinamento onde produz-se exclusivamente IgM; as células B fracamente autoreativas produzem grandes quantidades de IgD e tornam-se linfócitos B foliculares anérgicos (IgD++/IgM) com potencial para ativar a redenção clonal; e as células B sem autoreatividade desenvolvem-se em linfócitos B foliculares que produzem mais IgD que IgM (IgD+/IgM) ou em linfócitos da zona marginal que produzem mais IgM que IgD (IgM+/IgD).
Como mencionado anteriormente, os linfócitos da zona marginal irão produzir rapidamente anticorpos IgM de baixa afinidade para combater os agentes patogênicos, enquanto os linfócitos B foliculares irão formar centros germinativos para se diferenciarem em células plasmáticas e células B de memória que produzirão anticorpos de alta afinidade e especificidade aos antígenos.
Afinal de contas, por que precisamos de células dendríticas, linfócitos T e linfócitos B para desencadear a resposta humoral?
As células dendríticas não poderiam produzir os anticorpos? Ou mesmo os próprios linfócitos T auxiliares?
Como já fora mencionado anteriormente, as células dendríticas possuem uma coleção de receptores que lhes permitem reconhecer todo tipo de molécula invasora. Essas células são funcionais desde o nosso nascimento e, portanto, formam o elo entre o sistema imune inato e o sistema imune adaptativo.
De maneira simplificada, essas células exercem quatro principais funções: (1) detectar moléculas invasora através de seus receptores; (2) ingerir agentes patogênicos e fluído extracelular através da fagocitose e macropinocitose, respectivamente; (3) digerir moléculas invasoras e apresentar seus antígenos para os linfócitos T auxiliares; e (4) apresentar antígenos não-digeridos para os linfócitos B durante a maturação de afinidade de seus anticorpos nos centros germinativos (essa função é específica das células dendríticas foliculares) [1].
Se as células dendríticas fossem capazes de produzir anticorpos, não precisariam exercer a terceira e quarta função, correto?
Acontece que para exercer as duas primeiras funções, essas células precisam ter superfícies extensas. Isso faz com que o volume das células dendríticas seja 5–10 vezes maior que o dos linfócitos.
Indo direto ao ponto, as células dendríticas precisam produzir muito mais proteínas para exercer suas funções, o que faz com que sua proliferação (divisão celular) seja muito mais lenta.
Na verdade, elas sequer se dividem depois que atingem maturação, estando fadadas a exercer suas funções e morrer sem nunca deixar prole. Em contrapartida, os linfócitos podem se dividir em apenas 4 horas depois que são ativados e induzidos a proliferar.
Como vimos anteriormente, ao falar da Teoria de Seleção Clonal, os linfócitos B virgens que reconhecem um antígeno invasor são induzidos a proliferar nos centros germinativos, o que permite a maturação de afinidade e troca de classe de seus anticorpos.
Portanto, se dependêssemos da proliferação das células dendríticas para gerar anticorpos específicos a um vírus invasor, estaríamos fadados a perder a batalha.
Uma vez que é inviável a produção de anticorpos pelas células dendríticas, fica claro que essa função deve recair sobre outro tipo de célula, mas sob “supervisão” das células dendríticas, pois estas já são, afinal, especialistas em reconhecer moléculas invasoras.
Mas por que então essa função recai sobre os linfócitos B e não sobre os linfócitos T auxiliares, já que estes últimos interagem diretamente com as células dendríticas e produzem receptores similares aos anticorpos?
A resposta para essa pergunta é composta por dois motivos: a característica degenerada da estrutura de proteínas e a prevenção da autoimunidade.
Assim como o DNA, as proteínas são polímeros biológicos. Um polímero é formado pela união de duas ou mais unidades menores, chamadas monômeros.
No caso das proteínas, os monômeros são aminoácidos e temos ao todo 20 variedades deles. Todas as proteínas do seu corpo são formadas por apenas estes 20 aminoácidos, embora em diferentes combinações.
Por causa dessa característica, representamos as proteínas como uma “fila indiana” de aminoácidos ligados sequencialmente um ao outro. Não à toa, essa representação é chamada de sequência proteica.
Assim como acontece com os nucleotídeos na sequência de DNA, cada aminoácido da sequência proteica é representado por uma letra do alfabeto. A diferença em relação ao DNA é que ao invés de 4 letras (A, T, C ou G), podemos ter 20 letras diferentes na sequência proteica.
A sequência proteica é o que chamamos de estrutura primária da proteína, pois representa a relação mais básica entre os aminoácidos que a compõem: cada aminoácido interage apenas com o aminoácido que veio antes e aquele que o sucede na sequência.
Pois bem, mas o que isso tem a ver com os linfócitos e anticorpos?
Cada proteína tem uma sequência única, uma ordem específica na qual os aminoácidos foram ligados uns aos outros; um código de barras, por assim dizer.
A maior parte dos antígenos reconhecidos por anticorpos são proteínas, de maneira que eles reconhecem, na verdade, um pedaço dessas proteínas.
Portanto, para um anticorpo reconhecer uma proteína invasora (antígeno), bastaria que ele reconhecesse um pedaço singular de sua sequência proteica; mais especificamente, um pedaço da sequência cuja ordem dos aminoácidos só exista naquela proteína e em mais nenhuma outra.
O pedaço da proteína reconhecido pelo anticorpo é chamado de epítopo e nada mais é do que um grupo de aminoácidos em um pequeno espaço tridimensional. Ou seja, os anticorpos interagem com as proteínas por meio de contatos realizados em um espaço tridimensional.
Acontece que, ao contrário da estrutura primária (sequência proteica), as proteínas se dobram no espaço tridimensional para formar a estrutura proteica, também chamada de estrutura terciária.
É o formato tridimensional das proteínas que dão a elas suas funções; ou seja, é assim que elas são encontradas na natureza: dobradas. Uma consequência disso é que aminoácidos que estão muito distantes na sequencia proteica podem estar muito próximos na estrutura tridimensional.
Como os anticorpos reconhecem as proteínas por meio de contatos no espaço tridimensional, é bastante provável que os aminoácidos presentes na superfície de contato não sejam contíguos na sequencia proteica; isto é, não tem como garantir que eles sejam singulares para aquela proteína.
Uma outra consequência do dobramento de proteínas é que a estrutura proteica é degenerada. Brevemente, isso significa dizer que sequências proteicas diferentes podem dar origem a uma estrutura proteica similar.
Como vimos na Teoria da Seleção Clonal, a resposta primária é composta de anticorpos polireativos justamente para conseguir detectar formatos similares em antígenos diferentes, pois isso reduz enormemente a quantidade de linfócitos B que precisamos produzir para preencher todo espaço amostral de antígenos a serem detectados.
Ora, se os anticorpos precisam detectar o formato tridimensional dos antígenos (pois é assim que eles existem na natureza) e, ao mesmo tempo, precisam ser flexíveis o suficiente para detectar formatos similares, como então o sistema imune faz para garantir que eles não irão detectar formatos similares em nossas próprias proteínas? Entra em cena o linfócito T auxiliar.
Os linfócitos T auxiliares apresentam receptores que, embora parecidos com os anticorpos dos linfócitos B, são especializados em detectar fragmentos de 8–10 aminoácidos da sequência proteica. Ou seja, eles detectam um pedaço do “código de barras” da proteína invasora ao invés de reconhecer o formato tridimensional que elas possuem.
No entanto, para que isso aconteça, a estrutura terciaria da proteína precisa ser desfeita e fragmentos desdobrados precisam ser apresentados para os linfócitos T auxiliares.
É exatamente isso que as células dendríticas fazem ao ativar o linfócito T auxiliar e o mesmo que os linfócitos B virgens fazem quando solicitam a “confirmação” do antígeno a essas células (Figura 2B e 2C).
Após detectarem o formato tridimensional pertencente a um antígeno, os linfócitos B e as células dendríticas “ingerem” o antígeno e o degradam em pequenos fragmentos de 8–10 aminoácidos.
Esses fragmentos são então expostos na superfície celular por uma molécula especializada em apresenta-los em sua forma desdobrada: o Complexo Principal de Histocompatibilidade (MHC) de classe II. O MHC de classe II mantém os fragmentos linearizados (igual na sequencia proteica) para que sejam “lidos” pelos linfócitos T auxiliares.
Em conclusão, foi mais vantajoso para nosso sistema imune criar uma nova população de linfócitos especializados em detectar sequencias proteicas do que produzir uma quantidade muito maior de linfócitos B para detectar todos os antígenos com formatos tridimensionais específicos.
Essa dupla averiguação, uma por estrutura (mais flexível) e outra por sequência proteica (mais restrita), permite que o sistema imune tenha um controle maior sobre aquilo que é nosso daquilo que é invasor, sem ter que desperdiçar recursos a mais para isso. A natureza, além de sábia, é eficiente!
O que acontece quando somos vacinados ou infectados por um vírus?
Há uma ideia equivocada de que o único objetivo da vacinação contra a Covid-19 (ou qualquer outro patógeno) é que nosso corpo venha a produzir anticorpos IgG.
Embora a presença destes anticorpos no sangue confirme que nosso corpo produziu uma defesa de “longa duração” contra o vírus (coloquei entre aspas porque ainda não sabemos quanto tempo ela durará), a ausência deste anticorpo não significa que estamos desprotegidos.
Na realidade, não é a presença deste anticorpo que importa, mas sim a capacidade do nosso corpo em produzi-lo durante uma reinfecção.
A presença de IgG indica que nosso corpo está equipado com células plasmáticas capazes de produzir anticorpos para neutralizar especificamente partículas (vírions) do Sars-CoV-2 antes mesmo que estas consigam infectar nossas células.
As células plasmáticas de longa duração ficam hospedadas em nossa medula óssea e liberam constantemente pequenas quantidades de IgG e IgA de alta afinidade em nossa corrente sanguínea.
No entanto, o mesmo processo que dá origem às células plasmáticas também dá origem as células B de memória, que, embora não secretem IgG e IgA constitutivamente (o tempo todo), estão aptas a produzir grandes quantidades desses anticorpos assim que entram em contato com o antígeno viral.
As células B de memória são produzidas nos centros germinativos junto com as células plasmáticas e também produzem anticorpos de alta afinidade e especificidade ao antígeno.
A diferença é que enquanto as células plasmáticas se especializam na secreção de anticorpos, as células B de memória produzem principalmente anticorpos IgG, IgA ou IgE ancorados em suas membranas, as já mencionadas imunoglobulinas de membrana (mIg); essas células são conhecidas genericamente como células B de memória de classe trocada.
Há também uma pequena população de células B de memória, oriunda dos centros germinativos, que produzem quase que exclusivamente IgM de membrana (pois produz também um pouco de mIgD); estas células são conhecidas como células B de memória exclusivas em IgM (não confundir com células B de memória do tipo IgM, produzidas fora dos centros germinativos) e possuem cerca de 40% menos mutações em seus anticorpos que as células B de memória de classe trocada.
Isso significa que as células B de memória exclusivas em IgM permanecem menos tempo nos centros germinativos e, portanto, possuem anticorpos de menor afinidade que as células B de memória de classe trocada.
No entanto, como mencionado anteriormente, existem benefícios em se produzir IgM rapidamente, mesmo que este apresente menor afinidade.
O papel exercido pelas células B de memória é similar àquele exercido pelos linfócitos B virgens: elas se ligam aos antígenos através de suas mIgs e os apresentam para linfócitos T auxiliares, que por sua vez secretarão citocinas para ativar as células B de memória (Figura 2).
A diferença em relação aos linfócitos B virgens é que as células de memória já produzem anticorpos de alta afinidade e especificidade ao antígeno, de maneira que elas são muito mais eficientes na apresentação de antígenos aos linfócitos T auxiliares e, portanto, serão ativadas mais rapidamente.
Isso significa que a diferenciação de células B de memória em células plasmáticas (passando por um novo ciclo de maturação de afinidade), assim como a secreção de anticorpos específicos IgG e IgA, começa quase que simultaneamente à sua estimulação pelos antígenos.
A resposta humoral desencadeada pelos linfócitos B virgens é chamada de resposta primária, uma vez que acontece no primeiro contato com o antígeno, enquanto a resposta desencadeada pelas células B de memória é chamada de resposta secundária, pois acontece somente após o desenvolvimento da resposta primária.
A maior parte dos anticorpos produzidos durante a resposta secundária advém da ativação de células B de memória, e não das células plasmáticas de longa duração, de maneira que elas são nossa principal célula de defesa humoral durante uma reinfecção.
Agora que já sabemos como a resposta imune humoral se desenvolve, fica fácil entender que o sucesso de uma vacina está, na verdade, associado à sua capacidade em estimular principalmente a ativação e desenvolvimento dos linfócitos virgens ricos em IgD (IgD+/IgM); mais especificamente, os linfócitos B foliculares. Sem estes, não há a produção de anticorpos específicos de alta afinidade, pois são eles os precursores das células plasmáticas e células B de memória.
Como foi mencionado acima, as células B de memória precisam ser ativadas por linfócitos T auxiliares para secretarem anticorpos de alta afinidade, o que poderia atrasar a resposta secundária. Mas para isso há também as células T de memória auxiliares, que podem ativar as células B de memória sem a ajuda das células dendríticas.
Assim como há células T auxiliares, há também uma população de células T de memória citotóxicas, que atuam como sentinelas para destruir qualquer célula que venha a ser infectada pelo vírus.
Essas células citotóxicas são extremamente importantes e eficientes na defesa contra os vírus, mas não costumam ser levadas em conta na avaliação de eficiência das vacinas durante os ensaios clínicos.
Perceba que o contato inicial com o vírus produz um arsenal de células que estarão de prontidão para nos defender em caso de uma reinfecção.
Embora a presença de anticorpos específicos IgG (e IgA) no sangue seja extremamente relevante para neutralizar o vírus e impedir que este sequer penetre em nossas células, o aspecto mais importante da imunidade adaptativa é impedir que o vírus se multiplique a ponto de nos causar danos severos, o que também facilitará sua eliminação.
A forma como o sistema imune impede isso é preparando nosso corpo para desencadear uma resposta secundária quando encontrar o vírus novamente, que é muito mais robusta e eficiente que a resposta primária.
Hoje sabemos que mesmo os indivíduos que apresentam resultado negativo para anticorpos IgM e IgG possuem células T de memórias específicas contra o Sars-CoV-2; essas células foram encontradas tanto em hospedeiros assintomáticos quanto em indivíduos que apresentaram sintomas leves da Covid-19.
Em uma publicação anterior, eu expliquei a dinâmica de infecção de um vírus, usando como exemplo o vírus da Covid-19. Os parágrafos seguintes assumem que você sabe o que são as fases de latência, incubação, transmissão e convalescença.
Portanto, antes de continuar neste tópico, sugiro que você faça uma pausa e leia antes a referida publicação aqui (não precisa ler tudo, apenas o primeiro tópico), pois será necessário para o entendimento do restante do conteúdo.
Durante a primeira infeção pelo Sars-CoV-2, nosso sistema imune ainda não possui as melhores ferramentas para combater o vírus eficientemente. No entanto, muito embora seja o primeiro contato do nosso sistema imune com esse vírus, é provável que já tenhamos tido contato com vírus similares, de maneira que não estaremos inteiramente despreparados.
Se o vírus conseguir escapar do nosso sistema imune inato (ex. células dendríticas e macrófagos), poderá se replicar nos tecidos onde está melhor adaptado. Assim que este inicia efetivamente sua replicação, tem-se o fim da fase de latência e início da fase de transmissão viral.
No começo da fase de transmissão, a resposta imune adquirida pela infecção de outros vírus similares ao Sar-CoV-2 começa a atuar, ainda que de forma “amadora”.
De início, há principalmente produção de anticorpos IgM de baixa afinidade pelas células da zona marginal; enquanto isso os linfócitos B previamente ativados proliferam nos centros germinativos e refinam seus anticorpos.
Com sorte, esse arsenal prematuro de células de defesa será suficiente para desencadear uma reposta primária mais rápida, impedindo que o vírus cause danos aos nossos tecidos.
Para tal, é necessário a rápida produção de anticorpos IgG e IgA de alta afinidade. Se o sistema imune adaptativo obtiver sucesso, o indivíduo permanecerá assintomático durante todo o restante da fase de transmissão.
De fato, hoje sabemos que cerca de 20% das pessoas infectadas pelo Sar-Cov-2 permanecem assintomáticas porque conseguem desencadear uma resposta robusta ainda na fase de incubação.
Em geral, isso acontece porque essas pessoas foram previamente infectadas por outros coronavírus mais comuns, relacionados às gripes sazonais, e que lhe permitiram uma vantagem em relação ao Sars-CoV-2.
No entanto, para os outros 80% das pessoas, a resposta primária não será suficiente para manter o vírus em cheque (Figura 8; etapa 1).
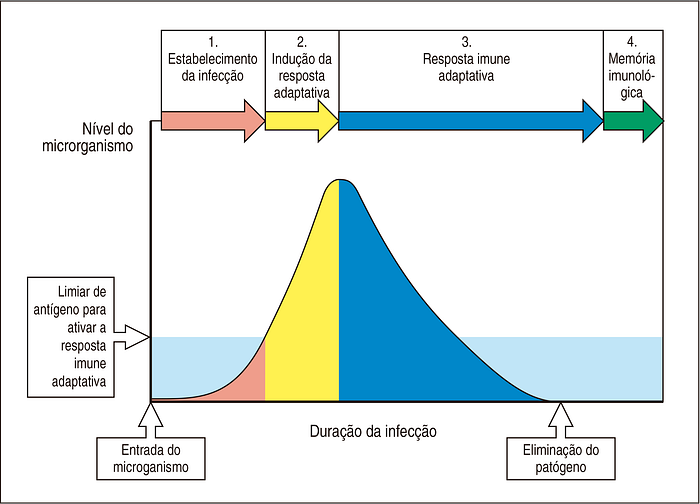
Uma vez que o vírus continua a se replicar, este começa a causar danos aos nossos tecidos e temos o surgimento dos primeiros sintomas (Figura 8; etapa 2). O dano causado pelo vírus atraí uma verdadeira “força-tarefa” de células de defesas para impedir que o mesmo continue a se replicar (resposta inflamatória).
A partir desse momento começam a aparecer as primeiras células plasmáticas e há liberação de anticorpos IgA e IgG de alta afinidade na corrente sanguínea (Figura 8; etapa 3). Esses anticorpos irão neutralizar o vírus e impedir a infecção de novas células.
Enquanto isso, os linfócitos T citotóxicos e as células do sistema imune inato irão remover as células previamente infectadas e as partículas virais já neutralizadas pelos anticorpos.
Anticorpos cada vez mais eficientes continuam ser produzidos e isso permite uma redução progressiva da carga viral até que o vírus seja totalmente eliminado (Figura 8; etapa 3).
Ao final, o sistema imune estará equipado com um novo arsenal de células plasmáticas, células B de memória e células T de memória (auxiliares e citotóxicas) para combater o vírus (Figura 8; etapa 4).
No entanto, para que todo esse arsenal seja produzido pelo sistema imune adaptativo, é necessário que a carga viral do hospedeiro alcance um certo limiar (Figura 8).
A velocidade com qual o vírus alcança esse limiar depende principalmente da carga viral inicial recebida pelo hospedeiro, bem como da resposta primária desencadeada pelo sistema imune.
Perceba que esses dois fatores podem variar muito de indivíduo para indivíduo, de maneira que o arsenal de células produzido pelo sistema imune adaptativo pode oscilar bastante ao final de infecções naturais. Ou seja, nem todos os indivíduos estarão protegidos igualmente.
Além disso, sabemos hoje que quanto maior a carga viral alcançada pelo vírus durante a infecção, maior é a quantidade de anticorpos IgG encontradas no sangue cerca de dois meses depois do surgimento dos primeiros sintomas.
No entanto, sabemos também que após esses dois meses acontece uma redução drástica de anticorpos IgG no sangue, igualmente proporcional à quantidade inicialmente produzida.
Portanto, quanto maior for a produção de anticorpos durante a infecção, maior é a redução dos mesmos nos meses seguintes. Isso significa que produzir mais anticorpos durante a infecção não necessariamente se traduz em uma proteção mais duradoura no caso da Covid-19.
Por fim, é muito importante ressaltar que uma infecção natural pelo vírus nem sempre termina com uma vitória do sistema imune adaptativo — além do óbvio risco de morte, há diversas pessoas que permanecem com sequelas decorrentes de danos causados pela Covid-19.
Em contrapartida, o que muda quando somos vacinados?
Em primeiro lugar, as doses aplicadas em vacinas são padronizadas para que induzam um efeito mínimo, de maneira que você diminui uma das variáveis mencionadas acima e induz uma resposta imune mais homogênea na população.
Em segundo lugar, as vacinas contra Covid-19 não se replicam no seu corpo e, portanto, não permitem o desenvolvimento da doença; o material entregue pela dose da vacina irá sempre diminuir com o passar do tempo, nunca aumentar.
Atualmente, as vacinas contra a Covid-19 entregam DNA (via vetor viral), RNA mensageiro, subpartículas virais ou partículas virais inativadas.
O objetivo de todas as vacinas é o mesmo: entregar uma amostra do coronavírus ao nosso sistema imune adaptativo para que ele possa induzir uma resposta secundária caso encontre o vírus no futuro.
E quais as desvantagens da vacina?
Na verdade, há apenas uma: a quantidade de material entregue em uma única dose da vacina pode não ser o suficiente para induzir o sistema imune adaptativo a produzir o arsenal de células necessário para desencadear uma resposta secundária.
Por esse motivo, pode ser necessário uma segunda dose da vacina, de maneira que pessoas vacinadas com apenas uma dose permanecem desprotegias até que a segunda dose faça efeito.
E por que algumas vacinas são mais eficientes que outras, podendo necessitar de apenas uma dose?
Vários fatores podem influenciar a efetividade de uma vacina, mas há três que costumam ser decisivos.
O primeiro, e mais óbvio, é o mecanismo de ação da vacina (veja o vídeo acima). Por exemplo, as vacinas de RNA e DNA entregam instruções para que as células do nosso corpo produzam uma proteína do Sars-CoV2, enquanto as vacinas de subpartículas e partículas inativadas entregam essas proteínas já prontas.
Com as instruções contidas no RNA e DNA, nossas células produzirão o tanto de proteína viral quanto forem capazes, o que em geral acaba sendo uma quantidade superior às proteínas prontas que são entregues pelas vacinas de subpartículas e partículas inativadas.
Ou seja, a quantidade de proteína viral produzida decorrente de vacinas de DNA e RNA é variável (e normalmente maior), enquanto que nas outras vacinas mencionadas a quantidade entregue é fixa (e normalmente menor). O que nos leva ao segundo fator: o método de entrega.
Independentemente do mecanismo de ação, a capacidade de uma vacina induzir uma resposta efetiva depende da eficiência com que ela faz chegar seu conteúdo para as células do sistema imune adaptativo.
Não existe um método de entrega que seja 100% eficiente, de forma que sempre haverá uma diferença entre aquilo que entra no seu braço por meio da vacina e aquilo que é efetivamente utilizado para estimular o sistema imune.
As vacinas da Janssen e AstraZeneca utilizam um vírus comum da gripe (adenovírus) para entregar DNA para nossas células; as vacinas da Pfizer e Moderna utilizam uma nanopartícula de gordura que se funde com nossas células para entregar moléculas de RNA; já a Coronavac utiliza uma versão inativada (morta) do próprio coronavírus para induzir nosso sistema imune.
O terceiro fator que influencia a efetividade de uma vacina é o uso de adjuvantes. Além de potencialmente fornecer uma quantidade insuficiente de proteína viral (antígeno) para estimular o sistema imune, as vacinas podem acabar por não estimular todas as células de defesa.
Como vimos anteriormente, o sistema imune adaptativo precisa da participação de várias células para conseguir montar uma resposta secundária, começando pelas células dendríticas. Acontece que alguns antígenos são melhores que outros para “chamar a atenção” de determinadas células de defesa.
Portanto, quando confeccionamos vacinas que focam em um determinado antígeno (as vacinas contra Covid-19 focam na proteína spike), nem sempre essa vacina consegue chamar a atenção de todas as células de defesa; dessa forma, adicionamos a essas vacinas moléculas complementares que irão ajudar a recrutar mais células de defesa para próximo do antígeno.
A Coronavac utliza sais de alumínio como adjuvantes; as vacinas da Moderna e Pfizer utilizam os lipídios presentes em suas nanopartículas de gordura como adjuvantes; e as vacinas da AstraZenica e Janssen utilizam um adenovírus como adjuvante.
Até momento, apenas a vacina da Janssen (subsidiaria de vacinas da Johnson & Johnson) consegue estimular uma resposta efetiva do sistema imune com apenas uma dose.
Mas então por que a vacina da AstraZenica, que utiliza o mesmo mecanismo de ação, método de entrega e adjuvante, precisa de duas doses?
Embora o processo de fabricação tenha influência no resultado final dessas vacinas (ex. concentração final de DNA por dose), é provável que o principal motivo seja a origem do adenovírus que foi utilizado como método de entrega.
Enquanto a AstraZenica utiliza um adenovírus que tipicamente infecta chipanzés, a Janssen utiliza um adenovírus que tipicamente infecta humanos.
Embora apenas a vacina da Janssen seja eficaz com a aplicação de uma única dose, um estudo recente mostrou que pessoas que testaram positivo para Covid-19 precisam de apenas uma dose da vacina da Pfizer para reativar a memória imunológica; a aplicação da segunda dose não traz benefício.
Isso permite com que todas as segundas doses dessas pessoas sejam redistribuídas para outras pessoas, acelerando o processo de vacinação.
Ainda sobre a eficiência das vacinas, é preciso esclarecer alguns pontos (veja o vídeo acima).
Assim como acontece no acompanhamento de uma epidemia, o dado mais fácil de se obter de uma população vacinada é referente ao início dos sintomas decorrentes da doença (veja aqui).
Quando escutamos que uma vacina contra Covid-19 possui eficiência 50, 70 ou 95%, essa informação refere-se exclusivamente à porcentagem de indivíduos vacinados que tiveram redução no surgimento de sintomas decorrentes da Covid-19.
Ou seja, uma eficiência de 70% indica que 7 a cada 10 pessoas vacinadas não apresentarão sintomas da Covid-19 mesmo que sejam infectadas pelo vírus.
Que sintomas são esses? Trata-se dos sintomas iniciais: dor de cabeça, coriza, tosse, dor de garganta, febre, perda de olfato, perda de paladar etc.
Como foi mencionado em um post anterior, o início dos sintomas marca o fim do período de incubação e indica que a multiplicação do vírus começou a causar os primeiros danos aos tecidos do hospedeiro; no caso da Covid-19, indica também que a carga viral está próxima do pico de transmissão (o hospedeiro tem maior probabilidade de transmitir o vírus).
Em resumo, quanto maior a eficiência de uma vacina, menor será a chance de indivíduos infectados apresentarem os primeiros sintomas e transmitirem o vírus para outros indivíduos.
Mas como, então, as vacinas impedem o surgimento dos primeiros sintomas?
Para isso ocorrer, o vírus deve ser impedido de se multiplicar, o que implica dizer que este deve ser neutralizado pelos anticorpos antes que consiga infectar novas células, bem como as células infectadas devem destruídas o mais rapidamente possível a fim de reduzir o tempo de multiplicação.
Como já vimos, a neutralização do vírus é decorrente da ação de anticorpos (Figura 1). Dessa forma, as vacinas mais eficientes na redução de sintomas são aquelas que induzem a secreção continua de anticorpos IgG na corrente sanguínea.
Tanto o surgimento de sintomas como a presença de anticorpos IgG são muito fáceis de se aferir e, portanto, são amplamente usados para medir a eficiência de uma vacina.
Mas seria este o melhor método para se averiguar a efetividade de uma vacina? A resposta é não; é apenas o mais fácil, tanto que este dado é obtido ainda nos ensaios clínicos.
Os sintomas por si só não são as principais consequências da doença; no caso da Covid-19, as principais consequências são a morte da população e a taxa de ocupação da UTIs, que por sua vez causa a morte de pessoas que precisam desse atendimento em decorrência de outras enfermidades.
E o que causa a morte e aumento da taxa de ocupação das UTIs?
A inabilidade do nosso corpo em controlar a infecção; ou seja, o prolongamento dos danos causados pelo vírus, e não o surgimento dos danos.
Embora todas as células de defesa contribuam para controlar a infecção, as células B de memória se destacam nessa função. São elas que irão produzir a grande maioria dos anticorpos de alta afinidade decorrentes de uma resposta secundária.
Essa alta produção de anticorpos demora um pouco para se concretizar e coincide justamente com o surgimento dos primeiros sintomas, de maneira que, embora o vírus cause danos aos tecidos, o sistema imune conseguirá rapidamente se livrar da infecção após esse evento.
Portanto, é a presença das células B de memória que efetivamente contribui para impedir o agravamento da doença, que por sua vez leva à morte e aumento da taxa de ocupação das UTIs.
Por esse motivo, a principal consequência da vacinação não é impedir o surgimento dos sintomas, mas sim diminuir a taxa de mortes e necessidade de tratamento intensivo pela população.
Essa informação não está disponível durante os ensaios clínicos, que são realizados em condições ideais e controladas, e só pode ser efetivamente aferida após a vacinação de uma grande parte da população em condições reais.
Logo, é superficial medir a efetividade de uma vacina apenas por sua eficiência em impedir o surgimento dos sintomas; o mais importante são os dados relativos à redução de casos graves e mortes decorrentes da doença.
De qualquer forma, é importante ressaltar que independentemente da eficiência da vacina (até mesmo se fosse 100%), esta não impede a transmissão do vírus!
Uma eficiência maior da vacina apenas diminui a probabilidade de transmissão do vírus, enquanto uma eficiência menor apenas indica que o hospedeiro terá maior probabilidade de ser acometido pelos sintomas iniciais da doença.
Mesmo que não haja surgimento dos sintomas, pessoas vacinadas, bem como não vacinadas, podem ser portadores assintomáticos da doença.
Desta forma, é primordial que pessoas vacinadas continuem a usar mascara e mantenham o distanciamento social para proteger o restante da população que ainda não foi vacinada.
Apenas quando estivermos todos vacinados é que será seguro interromper as medidas não farmacológicas de combate à Covid-19.
Por último, algumas pessoas têm dúvidas a respeito dos efeitos colaterais decorrentes da vacinação. Afinal, ter febre, tosse, dor de cabeça, diarreia etc., é um bom ou mau sinal?
Em geral, trata-se de um bom sinal, pois indica que seu corpo está induzindo uma resposta inflamatória, que nada mais é do que o recrutamento das suas células de defesa.
No entanto, assim com a ausência de anticorpos não indica ausência de uma resposta primária, a ausência de efeitos colaterais não indica a ausência de uma resposta inflamatória.
Esses tipos de consequências dependem da saúde e do sistema imune de cada indivíduo. Trata-se apenas de uma observação positiva; sua ausência não implica em um resultado negativo.
Dados recentes sobre a segurança das vacinas
Até o momento (janeiro de 2022), o único dado conclusivo em relação ao risco de aplicação das vacinas se resume a um aumento nos casos de miocardite (inflamção no coração) para jovens do sexo masculino (11 a 24 anos) até duas semanas após receber uma dose das vacinas de RNA mensageiro (Pfizer e Moderna). Esse aumento é mais significativo em homens de 18 anos e até 7 dias após o recebimento da segunda dose.
No entanto, não constataram mortes no estudo e embora quase todos os indivíduos com casos de miocardite tenham sido hospitalizados e monitorados clinicamente, eles experimentaram recuperação sintomática após receber apenas tratamento para dor.
Em contraste, os casos virais típicos de miocardite (não relacionados com a vacina) podem ter um curso clínico mais variável (que inclui mortes).
Constataram também que essa é a parcela da população americana que mais pratica esportes. Existe uma investigação em curso para saber se há correlação entre os casos de miocardite e a prática de esportes.
Por enquanto, a recomendação é que pessoas até 24 anos evitem tomar dose de vacinas que utilizem a tecnologia de RNA mensageiro (Pfizer e Moderna) e privilegiem as outras, como a CoronaVac e AstraZenica. Ademais, no caso de já ter tomado a dose de uma vacina de RNA mensageiro, este deve evitar praticar esportes até duas semanas após o recebimento da mesma.
 P
P
PDF
Como citar:
MARTINS-DE-SÁ, Diogo. A imunidade adquirida pela infecção natural de um vírus é melhor que imunidade adquirida pela vacina? Blog Diogo Martins de Sá. Brasília, 2 julho 2021, Disponível em: https://medium.com/p/6d61cb01e7cf/. Acesso em: dia mes ano.
Referências
1. Heesters, B. A., Myers, R. C., & Carroll, M. C. (2014). Follicular dendritic cells: dynamic antigen libraries. Nature reviews. Immunology, 14(7), 495–504. https://doi.org/10.1038/nri3689
2. Cyster J. G. (2010). B cell follicles and antigen encounters of the third kind. Nature immunology, 11(11), 989–996. https://doi.org/10.1038/ni.1946
3. Allen, C. D., & Cyster, J. G. (2008). Follicular dendritic cell networks of primary follicles and germinal centers: phenotype and function. Seminars in immunology, 20(1), 14–25. https://doi.org/10.1016/j.smim.2007.12.001
4. Gordon S. (2002). Pattern recognition receptors: doubling up for the innate immune response. Cell, 111(7), 927–930. https://doi.org/10.1016/s0092-8674(02)01201-1
5. Tian, R., Chen, M., Chai, S., Rong, X., Chen, B., Ren, W., Xu, S., & Yang, G. (2018). Divergent Selection of Pattern Recognition Receptors in Mammals with Different Ecological Characteristics. Journal of molecular evolution, 86(2), 138–149. https://doi.org/10.1007/s00239-018-9832-1
6. Zhang, Q., Zmasek, C. M., & Godzik, A. (2010). Domain architecture evolution of pattern-recognition receptors. Immunogenetics, 62(5), 263–272. https://doi.org/10.1007/s00251-010-0428-1
7. Marion J. (2014) Pattern Recognition Receptors: Evolution, Redundancy, and Cross Talk. In: Wells R., Bond J., Klinman J., Masters B., Bell E. (eds) Molecular Life Sciences. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-6436-5_369-1
8. Mellman I. (2013). Dendritic cells: master regulators of the immune response. Cancer immunology research, 1(3), 145–149. https://doi.org/10.1158/2326-6066.CIR-13-0102
9. Mbongue, J., Nicholas, D., Firek, A., & Langridge, W. (2014). The role of dendritic cells in tissue-specific autoimmunity. Journal of immunology research, 2014, 857143. https://doi.org/10.1155/2014/857143
10. Weiskopf, D., Schmitz, K. S., Raadsen, M. P., Grifoni, A., Okba, N., Endeman, H., van den Akker, J., Molenkamp, R., Koopmans, M., van Gorp, E., Haagmans, B. L., de Swart, R. L., Sette, A., & de Vries, R. D. (2020). Phenotype and kinetics of SARS-CoV-2-specific T cells in COVID-19 patients with acute respiratory distress syndrome. Science immunology, 5(48), eabd2071. https://doi.org/10.1126/sciimmunol.abd2071
11. Paces, J., Strizova, Z., Smrz, D., & Cerny, J. (2020). COVID-19 and the immune system. Physiological research, 69(3), 379–388. https://doi.org/10.33549/physiolres.934492
12. Sekine, T., Perez-Potti, A., Rivera-Ballesteros, O., Strålin, K., Gorin, J. B., Olsson, A., Llewellyn-Lacey, S., Kamal, H., Bogdanovic, G., Muschiol, S., Wullimann, D. J., Kammann, T., Emgård, J., Parrot, T., Folkesson, E., Karolinska COVID-19 Study Group, Rooyackers, O., Eriksson, L. I., Henter, J. I., Sönnerborg, A., … Buggert, M. (2020). Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell, 183(1), 158–168.e14. https://doi.org/10.1016/j.cell.2020.08.017
13. Newell, K. L., Clemmer, D. C., Cox, J. B., Kayode, Y. I., Zoccoli-Rodriguez, V., Taylor, H. E., Endy, T. P., Wilmore, J. R., & Winslow, G. M. (2021). Switched and unswitched memory B cells detected during SARS-CoV-2 convalescence correlate with limited symptom duration. PloS one, 16(1), e0244855. https://doi.org/10.1371/journal.pone.0244855
14. Mazzoni, A., Di Lauria, N., Maggi, L., Salvati, L., Vanni, A., Capone, M., Lamacchia, G., Mantengoli, E., Spinicci, M., Zammarchi, L., Kiros, S. T., Rocca, A., Lagi, F., Colao, M. G., Parronchi, P., Scaletti, C., Turco, L., Liotta, F., Rossolini, G. M., Cosmi, L., … COVID-19 Research Group (2021). First-dose mRNA vaccination is sufficient to reactivate immunological memory to SARS-CoV-2 in subjects who have recovered from COVID-19. The Journal of clinical investigation, 131(12), e149150. https://doi.org/10.1172/JCI149150
15. Cox, R. J., & Brokstad, K. A. (2020). Not just antibodies: B cells and T cells mediate immunity to COVID-19. Nature reviews. Immunology, 20(10), 581–582. https://doi.org/10.1038/s41577-020-00436-4
16. Zhao, J., Yuan, Q., Wang, H., Liu, W., Liao, X., Su, Y., Wang, X., Yuan, J., Li, T., Li, J., Qian, S., Hong, C., Wang, F., Liu, Y., Wang, Z., He, Q., Li, Z., He, B., Zhang, T., Fu, Y., … Zhang, Z. (2020). Antibody Responses to SARS-CoV-2 in Patients With Novel Coronavirus Disease 2019. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America, 71(16), 2027–2034. https://doi.org/10.1093/cid/ciaa344
17. Zhou, W., Xu, X., Chang, Z., Wang, H., Zhong, X., Tong, X., Liu, T., & Li, Y. (2021). The dynamic changes of serum IgM and IgG against SARS-CoV-2 in patients with COVID-19. Journal of medical virology, 93(2), 924–933. https://doi.org/10.1002/jmv.26353
18. Quast, I., & Tarlinton, D. (2021). B cell memory: understanding COVID-19. Immunity, 54(2), 205–210. https://doi.org/10.1016/j.immuni.2021.01.014
19. Pardi, N., Hogan, M. J., Porter, F. W., & Weissman, D. (2018). mRNA vaccines — a new era in vaccinology. Nature reviews. Drug discovery, 17(4), 261–279. https://doi.org/10.1038/nrd.2017.243
20. Gutierrez, L., Beckford, J., & Alachkar, H. (2020). Deciphering the TCR Repertoire to Solve the COVID-19 Mystery. Trends in pharmacological sciences, 41(8), 518–530. https://doi.org/10.1016/j.tips.2020.06.001
21. Zhang, Y., Zeng, G., Pan, H., Li, C., Hu, Y., Chu, K., Han, W., Chen, Z., Tang, R., Yin, W., Chen, X., Hu, Y., Liu, X., Jiang, C., Li, J., Yang, M., Song, Y., Wang, X., Gao, Q., & Zhu, F. (2021). Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18–59 years: a randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. The Lancet. Infectious diseases, 21(2), 181–192. https://doi.org/10.1016/S1473-3099(20)30843-4
22. Silva-Cayetano, A., Foster, W. S., Innocentin, S., Belij-Rammerstorfer, S., Spencer, A. J., Burton, O. T., Fra-Bidó, S., Le Lee, J., Thakur, N., Conceicao, C., Wright, D., Barrett, J., Evans-Bailey, N., Noble, C., Bailey, D., Liston, A., Gilbert, S. C., Lambe, T., & Linterman, M. A. (2021). A booster dose enhances immunogenicity of the COVID-19 vaccine candidate ChAdOx1 nCoV-19 in aged mice. Med (New York, N.Y.), 2(3), 243–262.e8. https://doi.org/10.1016/j.medj.2020.12.006
23. Silva-Cayetano, A., Foster, W. S., Innocentin, S., Belij-Rammerstorfer, S., Spencer, A. J., Burton, O. T., Fra-Bidó, S., Le Lee, J., Thakur, N., Conceicao, C., Wright, D., Barrett, J., Evans-Bailey, N., Noble, C., Bailey, D., Liston, A., Gilbert, S. C., Lambe, T., & Linterman, M. A. (2021). A booster dose enhances immunogenicity of the COVID-19 vaccine candidate ChAdOx1 nCoV-19 in aged mice. Med (New York, N.Y.), 2(3), 243–262.e8. https://doi.org/10.1016/j.medj.2020.12.006
24. Hendaus, M. A., & Jomha, F. A. (2021). mRNA Vaccines for COVID-19: A Simple Explanation. Qatar medical journal, 2021(1), 07. https://doi.org/10.5339/qmj.2021.07
25. Long, Q. X., Tang, X. J., Shi, Q. L., Li, Q., Deng, H. J., Yuan, J., Hu, J. L., Xu, W., Zhang, Y., Lv, F. J., Su, K., Zhang, F., Gong, J., Wu, B., Liu, X. M., Li, J. J., Qiu, J. F., Chen, J., & Huang, A. L. (2020). Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nature medicine, 26(8), 1200–1204. https://doi.org/10.1038/s41591-020-0965-6
26. Bettini, E., & Locci, M. (2021). SARS-CoV-2 mRNA Vaccines: Immunological Mechanism and Beyond. Vaccines, 9(2), 147. https://doi.org/10.3390/vaccines9020147
27. Yang, S., Jerome, K. R., Greninger, A. L., Schiffer, J. T., & Goyal, A. (2021). Endogenously Produced SARS-CoV-2 Specific IgG Antibodies May Have a Limited Impact on Clearing Nasal Shedding of Virus during Primary Infection in Humans. Viruses, 13(3), 516. https://doi.org/10.3390/v13030516
28. Carsetti, R., Köhler, G., & Lamers, M. C. (1993). A role for immunoglobulin D: interference with tolerance induction. European journal of immunology, 23(1), 168–178. https://doi.org/10.1002/eji.1830230127
29. Chen, K., & Cerutti, A. (2011). The function and regulation of immunoglobulin D. Current opinion in immunology, 23(3), 345–352. https://doi.org/10.1016/j.coi.2011.01.006
30. Übelhart, R., Hug, E., Bach, M. P., Wossning, T., Dühren-von Minden, M., Horn, A. H., Tsiantoulas, D., Kometani, K., Kurosaki, T., Binder, C. J., Sticht, H., Nitschke, L., Reth, M., & Jumaa, H. (2015). Responsiveness of B cells is regulated by the hinge region of IgD. Nature immunology, 16(5), 534–543. https://doi.org/10.1038/ni.3141
31. Gutzeit, C., Chen, K., & Cerutti, A. (2018). The enigmatic function of IgD: some answers at last. European journal of immunology, 48(7), 1101–1113. https://doi.org/10.1002/eji.201646547
32. Edholm, E. S., Bengten, E., & Wilson, M. (2011). Insights into the function of IgD. Developmental and comparative immunology, 35(12), 1309–1316. https://doi.org/10.1016/j.dci.2011.03.002
33. Kato, Y., Abbott, R. K., Freeman, B. L., Haupt, S., Groschel, B., Silva, M., Menis, S., Irvine, D. J., Schief, W. R., & Crotty, S. (2020). Multifaceted Effects of Antigen Valency on B Cell Response Composition and Differentiation In Vivo. Immunity, 53(3), 548–563.e8. https://doi.org/10.1016/j.immuni.2020.08.001
34. Rosenspire, A. J., & Chen, K. (2015). Anergic B Cells: Precarious On-Call Warriors at the Nexus of Autoimmunity and False-Flagged Pathogens. Frontiers in immunology, 6, 580. https://doi.org/10.3389/fimmu.2015.00580
35. Zikherman, J., Parameswaran, R., & Weiss, A. (2012). Endogenous antigen tunes the responsiveness of naive B cells but not T cells. Nature, 489(7414), 160–164. https://doi.org/10.1038/nature11311
36. Minguet, S., Dopfer, E. P., & Schamel, W. W. (2010). Low-valency, but not monovalent, antigens trigger the B-cell antigen receptor (BCR). International immunology, 22(3), 205–212. https://doi.org/10.1093/intimm/dxp129
37. Duty, J. A., Szodoray, P., Zheng, N. Y., Koelsch, K. A., Zhang, Q., Swiatkowski, M., Mathias, M., Garman, L., Helms, C., Nakken, B., Smith, K., Farris, A. D., & Wilson, P. C. (2009). Functional anergy in a subpopulation of naive B cells from healthy humans that express autoreactive immunoglobulin receptors. The Journal of experimental medicine, 206(1), 139–151. https://doi.org/10.1084/jem.20080611
38. Sabouri, Z., Perotti, S., Spierings, E., Humburg, P., Yabas, M., Bergmann, H., Horikawa, K., Roots, C., Lambe, S., Young, C., Andrews, T. D., Field, M., Enders, A., Reed, J. H., & Goodnow, C. C. (2016). IgD attenuates the IgM-induced anergy response in transitional and mature B cells. Nature communications, 7, 13381. https://doi.org/10.1038/ncomms13381
39. Brink, R., Goodnow, C. C., Crosbie, J., Adams, E., Eris, J., Mason, D. Y., Hartley, S. B., & Basten, A. (1992). Immunoglobulin M and D antigen receptors are both capable of mediating B lymphocyte activation, deletion, or anergy after interaction with specific antigen. The Journal of experimental medicine, 176(4), 991–1005. https://doi.org/10.1084/jem.176.4.991
40. Tan, C., Noviski, M., Huizar, J., & Zikherman, J. (2019). Self-reactivity on a spectrum: A sliding scale of peripheral B cell tolerance. Immunological reviews, 292(1), 37–60. https://doi.org/10.1111/imr.12818
41. Burnett, D. L., Reed, J. H., Christ, D., & Goodnow, C. C. (2019). Clonal redemption and clonal anergy as mechanisms to balance B cell tolerance and immunity. Immunological reviews, 292(1), 61–75. https://doi.org/10.1111/imr.12808
42. Sabouri, Z., Schofield, P., Horikawa, K., Spierings, E., Kipling, D., Randall, K. L., Langley, D., Roome, B., Vazquez-Lombardi, R., Rouet, R., Hermes, J., Chan, T. D., Brink, R., Dunn-Walters, D. K., Christ, D., & Goodnow, C. C. (2014). Redemption of autoantibodies on anergic B cells by variable-region glycosylation and mutation away from self-reactivity. Proceedings of the National Academy of Sciences of the United States of America, 111(25), E2567–E2575. https://doi.org/10.1073/pnas.1406974111
43. Noviski, M., & Zikherman, J. (2018). Control of autoreactive B cells by IgM and IgD B cell receptors: maintaining a fine balance. Current opinion in immunology, 55, 67–74. https://doi.org/10.1016/j.coi.2018.09.015
44. Quách, T. D., Manjarrez-Orduño, N., Adlowitz, D. G., Silver, L., Yang, H., Wei, C., Milner, E. C., & Sanz, I. (2011). Anergic responses characterize a large fraction of human autoreactive naive B cells expressing low levels of surface IgM. Journal of immunology (Baltimore, Md. : 1950), 186(8), 4640–4648. https://doi.org/10.4049/jimmunol.1001946
45. Pone, E. J., Zan, H., Zhang, J., Al-Qahtani, A., Xu, Z., & Casali, P. (2010). Toll-like receptors and B-cell receptors synergize to induce immunoglobulin class-switch DNA recombination: relevance to microbial antibody responses. Critical reviews in immunology, 30(1), 1–29. https://doi.org/10.1615/critrevimmunol.v30.i1.10
46. Pone, E. J., Xu, Z., White, C. A., Zan, H., & Casali, P. (2012). B cell TLRs and induction of immunoglobulin class-switch DNA recombination. Frontiers in bioscience (Landmark edition), 17, 2594–2615. https://doi.org/10.2741/4073
47. Hua, Z., & Hou, B. (2013). TLR signaling in B-cell development and activation. Cellular & molecular immunology, 10(2), 103–106. https://doi.org/10.1038/cmi.2012.61
48. Suthers, A. N., & Sarantopoulos, S. (2017). TLR7/TLR9- and B Cell Receptor-Signaling Crosstalk: Promotion of Potentially Dangerous B Cells. Frontiers in immunology, 8, 775. https://doi.org/10.3389/fimmu.2017.00775
49. Li, Y., Li, H., & Weigert, M. (2002). Autoreactive B cells in the marginal zone that express dual receptors. The Journal of experimental medicine, 195(2), 181–188. https://doi.org/10.1084/jem.20011453
50. Daly, C. A., Spurrier, M. A., Jennings-Gee, J. E., & Haas, K. M. (2020). B Cell Subsets Differentially Contribute to the T Cell-Independent Memory Pool. Journal of immunology (Baltimore, Md. : 1950), 205(9), 2362–2374. https://doi.org/10.4049/jimmunol.1901453
51. Seifert, M., & Küppers, R. (2009). Molecular footprints of a germinal center derivation of human IgM+(IgD+)CD27+ B cells and the dynamics of memory B cell generation. The Journal of experimental medicine, 206(12), 2659–2669. https://doi.org/10.1084/jem.20091087
52. Bautista, D., Vásquez, C., Ayala-Ramírez, P., Téllez-Sosa, J., Godoy-Lozano, E., Martínez-Barnetche, J., Franco, M., & Angel, J. (2020). Differential Expression of IgM and IgD Discriminates Two Subpopulations of Human Circulating IgM+IgD+CD27+ B Cells That Differ Phenotypically, Functionally, and Genetically. Frontiers in immunology, 11, 736. https://doi.org/10.3389/fimmu.2020.00736
53. Capolunghi, F., Rosado, M. M., Sinibaldi, M., Aranburu, A., & Carsetti, R. (2013). Why do we need IgM memory B cells?. Immunology letters, 152(2), 114–120. https://doi.org/10.1016/j.imlet.2013.04.007
54. Zhang X. (2013). Regulatory functions of innate-like B cells. Cellular & molecular immunology, 10(2), 113–121. https://doi.org/10.1038/cmi.2012.63
55. Marinkovic, D., & Marinkovic, T. (2020). Putative role of marginal zone B cells in pathophysiological processes. Scandinavian journal of immunology, 92(3), e12920. https://doi.org/10.1111/sji.12920
56. Martin, F., Oliver, A. M., & Kearney, J. F. (2001). Marginal zone and B1 B cells unite in the early response against T-independent blood-borne particulate antigens. Immunity, 14(5), 617–629. https://doi.org/10.1016/s1074-7613(01)00129-7
57. Martin, F., & Kearney, J. F. (2002). Marginal-zone B cells. Nature reviews. Immunology, 2(5), 323–335. https://doi.org/10.1038/nri799
58. Fink K. (2012). Origin and Function of Circulating Plasmablasts during Acute Viral Infections. Frontiers in immunology, 3, 78. https://doi.org/10.3389/fimmu.2012.00078
59. Lechner, M., Engleitner, T., Babushku, T., Schmidt-Supprian, M., Rad, R., Strobl, L. J., & Zimber-Strobl, U. (2021). Notch2-mediated plasticity between marginal zone and follicular B cells. Nature communications, 12(1), 1111. https://doi.org/10.1038/s41467-021-21359-1
60. Allman, D., & Pillai, S. (2008). Peripheral B cell subsets. Current opinion in immunology, 20(2), 149–157. https://doi.org/10.1016/j.coi.2008.03.014
61. Graf, R., Seagal, J., Otipoby, K. L., Lam, K. P., Ayoub, S., Zhang, B., Sander, S., Chu, V. T., & Rajewsky, K. (2019). BCR-dependent lineage plasticity in mature B cells. Science (New York, N.Y.), 363(6428), 748–753. https://doi.org/10.1126/science.aau8475
62. Pillai, S., & Cariappa, A. (2009). The follicular versus marginal zone B lymphocyte cell fate decision. Nature reviews. Immunology, 9(11), 767–777. https://doi.org/10.1038/nri2656
63. Ellebedy, A. H., Jackson, K. J., Kissick, H. T., Nakaya, H. I., Davis, C. W., Roskin, K. M., McElroy, A. K., Oshansky, C. M., Elbein, R., Thomas, S., Lyon, G. M., Spiropoulou, C. F., Mehta, A. K., Thomas, P. G., Boyd, S. D., & Ahmed, R. (2016). Defining antigen-specific plasmablast and memory B cell subsets in human blood after viral infection or vaccination. Nature immunology, 17(10), 1226–1234. https://doi.org/10.1038/ni.3533
64. Wen, L., Brill-Dashoff, J., Shinton, S. A., Asano, M., Hardy, R. R., & Hayakawa, K. (2005). Evidence of marginal-zone B cell-positive selection in spleen. Immunity, 23(3), 297–308. https://doi.org/10.1016/j.immuni.2005.08.007
65. Myers, D. R., Zikherman, J., & Roose, J. P. (2017). Tonic Signals: Why Do Lymphocytes Bother?. Trends in immunology, 38(11), 844–857. https://doi.org/10.1016/j.it.2017.06.010
66. Cyster, J. G., & Allen, C. (2019). B Cell Responses: Cell Interaction Dynamics and Decisions. Cell, 177(3), 524–540. https://doi.org/10.1016/j.cell.2019.03.016
67. Zabel, F., Fettelschoss, A., Vogel, M., Johansen, P., Kündig, T. M., & Bachmann, M. F. (2017). Distinct T helper cell dependence of memory B-cell proliferation versus plasma cell differentiation. Immunology, 150(3), 329–342. https://doi.org/10.1111/imm.12688
68. Radbruch, A., Muehlinghaus, G., Luger, E. O., Inamine, A., Smith, K. G., Dörner, T., & Hiepe, F. (2006). Competence and competition: the challenge of becoming a long-lived plasma cell. Nature reviews. Immunology, 6(10), 741–750. https://doi.org/10.1038/nri1886
69. Gao, M., & Skolnick, J. (2010). Structural space of protein-protein interfaces is degenerate, close to complete, and highly connected. Proceedings of the National Academy of Sciences of the United States of America, 107(52), 22517–22522. https://doi.org/10.1073/pnas.1012820107
70. Xu, J. L., & Davis, M. M. (2000). Diversity in the CDR3 region of V(H) is sufficient for most antibody specificities. Immunity, 13(1), 37–45. https://doi.org/10.1016/s1074-7613(00)00006-6
71. Akbar, R., Robert, P. A., Pavlović, M., Jeliazkov, J. R., Snapkov, I., Slabodkin, A., Weber, C. R., Scheffer, L., Miho, E., Haff, I. H., Haug, D., Lund-Johansen, F., Safonova, Y., Sandve, G. K., & Greiff, V. (2021). A compact vocabulary of paratope-epitope interactions enables predictability of antibody-antigen binding. Cell reports, 34(11), 108856. https://doi.org/10.1016/j.celrep.2021.108856
72. Davis M. M. (2004). The evolutionary and structural ‘logic’ of antigen receptor diversity. Seminars in immunology, 16(4), 239–243. https://doi.org/10.1016/j.smim.2004.08.003
73. Rees A. R. (2020). Understanding the human antibody repertoire. mAbs, 12(1), 1729683. https://doi.org/10.1080/19420862.2020.1729683
74. Perelson, A. S., & Oster, G. F. (1979). Theoretical studies of clonal selection: minimal antibody repertoire size and reliability of self-non-self discrimination. Journal of theoretical biology, 81(4), 645–670. https://doi.org/10.1016/0022-5193(79)90275-3
75. Hong, B., Wu, Y., Li, W., Wang, X., Wen, Y., Jiang, S., Dimitrov, D. S., & Ying, T. (2018). In-Depth Analysis of Human Neonatal and Adult IgM Antibody Repertoires. Frontiers in immunology, 9, 128. https://doi.org/10.3389/fimmu.2018.00128
76. de Greef, P. C., Oakes, T., Gerritsen, B., Ismail, M., Heather, J. M., Hermsen, R., Chain, B., & de Boer, R. J. (2020). The naive T-cell receptor repertoire has an extremely broad distribution of clone sizes. eLife, 9, e49900. https://doi.org/10.7554/eLife.49900
77. De Boer, R. J., & Perelson, A. S. (1993). How diverse should the immune system be?. Proceedings. Biological sciences, 252(1335), 171–175. https://doi.org/10.1098/rspb.1993.0062
78. Soto, C., Bombardi, R. G., Branchizio, A., Kose, N., Matta, P., Sevy, A. M., Sinkovits, R. S., Gilchuk, P., Finn, J. A., & Crowe, J. E., Jr (2019). High frequency of shared clonotypes in human B cell receptor repertoires. Nature, 566(7744), 398–402. https://doi.org/10.1038/s41586-019-0934-8
79. Briney, B., Inderbitzin, A., Joyce, C., & Burton, D. R. (2019). Commonality despite exceptional diversity in the baseline human antibody repertoire. Nature, 566(7744), 393–397. https://doi.org/10.1038/s41586-019-0879-y
80. Oster, Matthew E et al. “Myocarditis Cases Reported After mRNA-Based COVID-19 Vaccination in the US From December 2020 to August 2021.” JAMA vol. 327,4 (2022): 331–340. https://doi.org/10.1001/jama.2021.24110
